Ацетабулярия
| Ацетабулярия | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | ||||||||||||||
| Научная классификация | ||||||||||||||
|
Домен: Царство: Подцарство: Отдел: Класс: Порядок: Семейство: Род: Ацетабулярия |
||||||||||||||
| Международное научное название | ||||||||||||||
| Acetabularia J.V.Lamour., 1812 | ||||||||||||||
| Виды | ||||||||||||||
|
См. текст |
||||||||||||||
| ||||||||||||||
Ацетабуля́рия (лат. Acetabularia), также известная как «бокал русалки»[1] — род зелёных водорослей[2], гигантская сифоновая одноклеточная водоросль[3] с единственным клеточным ядром[4], расположенным в нижней части организма.
Известны и другие клетки больших размеров — нервные клетки (например, гигантские аксоны кальмара),[5][6] лубяные волокна растений или яйца птиц, пресмыкающихся и первозверей. Но в данном случае размера порядка нескольких сантиметров достигает взрослый одноклеточный организм. Впервые это качество ацетабулярии описал в 1930-е годы немецкий биолог Иоахим Геммерлинг[4][7].
Стебелёк взрослого растения имеет длину от 2—3 см[8] до 4—6 см, а шляпка (зонтик) — до 1 см в диаметре[9]. Для отдельных видов длина стебелька достигает 10 см[10] и даже 18 см[11].
Ацетабулярия обитает в морях в условиях тропического и субтропического климата[2] (в частности, в Средиземном море, в бассейне Индийского океана и Карибского моря)[4].
Ацетабулярия живёт на мелководье и часто повреждается прибоем. Эволюционное приспособление к этой среде — способность к регенерации всех утраченных частей, кроме клеточного ядра. При этом единственное ядро этого одноклеточного растения находится в ризоиде (ножке), прикреплённой к камням[4].
В лабораторных условиях используется как удобный объект при исследовании ядерно-плазматического взаимодействия[2].
Первые публикации[править | править код]
Организм был впервые описан в 1586 году (Mattoli, 1586), через 54 года последовала вторая публикация (Parkinson, 1640) и 172 года позднее — третья (Linneaus, 1758). В 1930-е гг. Геммерлинг открыл одноклеточный характер водоросли. Взрывной рост публикаций последовал между 1950 и 1980 годами, достигая пяти сотен за десятилетие (в период наибольшего подъёма в 1970-е — главным образом, по тематике ритмов и электрофизиологии), после чего он пошёл на спад (график числа публикаций приведён по ссылке)[12].
Классификация[править | править код]
Согласно классификации, принятой в СССР (по данным В. И. Кефели, 1978), Ацетабулярия (Acetabularia) принадлежит к типу зелёных водорослей (Chlorophyta), к классу равножгутиковых (Isocontae), порядку сифоновых (Siphonales), семейству дазикладовых (Dasycladaceae)[10].
Дазикладовые (Dasycladaceae) теперь классифицируются как член класса Ульвофициевые (Ulvophyceae) (Graham et Wilcox, 2000), хотя van den Hoek и др. (1993), согласно определённой ими комбинации свойств, размещали их отдельный класс — Dasycladophyceae[13].
Acetabularia также является синонимом одного из родов грибов — Cyphellopus (Index Fungorum, MycoBank).
Виды[править | править код]
- Acetabularia acetabulum
- Acetabularia antillana
- Acetabularia calyculus
- Acetabularia crenulata
- Acetabularia dentata
- Acetabularia farlowii
- Acetabularia kilneri
- Acetabularia major
- Acetabularia myriospora
- Acetabularia peniculus
- Acetabularia ryukyuensis
- Acetabularia schenkii
- Acetabularia toxasii
Строение[править | править код]

Ацетабулярия представляет собой одноклеточный организм[14], гигантскую одноядерную клетку, имеющую сложное строение[15].
Состоит из ризоида с ядром, стебелька и зонтика (шапочки)[3].
Стебелёк, увенчанный сверху шапочкой, имеет длину до 5 см. Внизу стебелька находится ризоид, где расположено единственное у этого одноклеточного организма ядро[15].
Ацетабулярия живёт на мелководье. Ножкой (ризоидом) она прикрепляется к каменистому грунту[16].
Шапочка (шляпка) образована мутовкой сросшихся или несросшихся гаметангиев[2]. Так, у средиземноморской A. mediterranea гаметангии срослись в единую шляпку или зонтик[17]. У другого вида — А. crenulata из Карибского моря — дольки (гаметангии) зонтика имеют форму банана[18] и радиально расходятся от вершины стебелька[11].
Жизненный цикл[править | править код]

В природе полный жизненный цикл Aceiabularia mediterranea составляет около трёх лет, а в лабораторных условиях может быть сокращён до нескольких месяцев[9] (примерно до 6 месяцев)[2].
1 — Из прошедшей зимовку цисты весной выходят жгутиковые клетки (гаметы) с мягкой оболочкой и двумя жгутиками на конце[2][9].
2 — Гаметы некоторое время плавают, а затем попарно копулируют[2][9]. При этом пары составляют гаметы из разных цист.[4]:96
3 — Примерно через два дня образуется одна диплоидная клетка (зигота)[9].
4 — На первом году жизни клетка состоит из ризоида (разветвлённой ножки), которой она крепится к субстрату, и стебелька без зонтика. Осенью стебелёк засыхает и отваливается, и оставшийся ризоид зимует за счёт запасённых веществ[2][9].
5 — Следующей весной вырастает новый стебелёк с зачатком зонтика (одна или несколько стерильных мутовок, которые осенью отмирают)[2][9].
6 — На третий год формируются зрелые стебелёк и зонтик (мутовка из гаметангиев). В период завершения вегетативной фазы ядро многократно делится (распадается)[11], и образованные мелкие вторичные ядра (10—20 тысяч) перемещаются в зонтик (в гаметангии)[2][9][10].
7 — Ядра собираются в цисты с толстыми стенками. Осенью цисты выходят из зонтика и зимуют в свободном состоянии[2][9]. Согласно одной точке зрения, внутри цисты осуществляется несколько ядерных делений, последнее из которых носит редукционный характер (мейоз). Согласно другой, более современной[19] точке зрения, редукционное деление производится на более ранней стадии — при распаде гигантского ядра, расположенного в ризоиде[11]. Вокруг каждого ядра цисты концентрируется цитоплазма и формируется клеточная мембрана[2][9].
Существует точка зрения (Berger, Liddle, 2003), согласно которой 3-летний жизненный цикл средиземноморской ацетабулярии A. mediterranea, воспроизводимый учебной литературой из источников XIX века, на самом деле не подтвержден наблюдениями и опытами. Эксперименты этих исследователей показали, что, хотя клетка может впадать в дремлющее состояние в холодной 10 °C воде, она также может завершить свой жизненный цикл в течение одного сезона[20].
Выращивание в искусственной среде[править | править код]
Впервые технику выращивания ацетабулярии описали Геммерлинг (1931) и Beth (1953)[21].
Ацетабулярия при переносе её в лабораторию отказывалась расти даже в морской воде из соответствующих мест обитания, пока Геммерлинг не добавил в воду отвар садовой земли[22].
Состав среды для выращивания Acetabularia mediterranea, который применялся длительное время в различных лабораториях[23]:
| Компоненты | Частей на литр |
|---|---|
| NaNO3 | 100 мг |
| Na2HPO4 | 20 мг |
| Экстракт почвы | 2—5 мл |
| Натуральная морская вода | 1 л |
Впоследствии Шепардом (1970) была разработана полностью искусственная среда на основе дистиллированной воды, нескольких различных солей с добавлением витаминов (таблица состава приведена по ссылке), которая позволяет поддерживать нормальное развитие ацетабулярии[21].
Регенерация[править | править код]
Ацетабулярия обладает особенностью, которая позволяет ей синтезировать свой зонтик и другие части после удаления корешка или расположенного в корешке ядра[24].
Энуклеированное (лишённое ядра) растение способно восстановить утраченные части: зонтик, ризоид: всё, за исключением ядра. Такие растения погибают через несколько месяцев. Напротив, части этого одноклеточного растения с ядром способны неоднократно восстанавливаться после повреждения[9].
Производились следующие опыты с регенерацией ацетабулярии[25] [26] [27]:
-
 Ампутация ножки (ризоида), которая содержит единственное клеточное ядро растения. Образуется новый ризоид, который, однако, не имеет ядра. Клетка может выжить в благоприятных условиях несколько месяцев, но уже не способна к размножению
Ампутация ножки (ризоида), которая содержит единственное клеточное ядро растения. Образуется новый ризоид, который, однако, не имеет ядра. Клетка может выжить в благоприятных условиях несколько месяцев, но уже не способна к размножению -
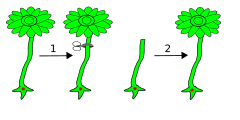 Удаление зонтика (шляпки). После этого одноклеточное растение формирует новую шляпку
Удаление зонтика (шляпки). После этого одноклеточное растение формирует новую шляпку -
 Стебелёк отрезан чуть выше ризоида. Это приводит к образованию второго ризоида
Стебелёк отрезан чуть выше ризоида. Это приводит к образованию второго ризоида -
 Удаление ризоида и шляпки (оставлена центральная часть). Происходит их восстановление (без ядра)
Удаление ризоида и шляпки (оставлена центральная часть). Происходит их восстановление (без ядра) -
 Стебелёк отрезан ближе к шляпке. Это приводит к образованию второй шляпки
Стебелёк отрезан ближе к шляпке. Это приводит к образованию второй шляпки -
 Удаление сначала шляпки, а затем - ризоида с ядром. Из этого эксперимента было показано, что ядро отвечает за обучение
Удаление сначала шляпки, а затем - ризоида с ядром. Из этого эксперимента было показано, что ядро отвечает за обучение






В России опытами по регенерации ацетабулярии занимался академик РАН Лев Сандахчиев[28].
Трансплантация ядра[править | править код]

Ядро ацетабулярии отличается жизнеспособностью: будучи выделенным из организма и очищенным, оно может быть сохранено в растворе сахара в течение 24 часов[4].
При пересадке в ацетабулярию ядра другого вида новое ядро «отдаёт команды» по постройке новой шляпки. Но если в стебле ещё остался запас «шляпкообразующего вещества» старого типа, то в результате образуется смешанная по своим свойствам клетка-гибрид[4].
Геммерлинг производил замещение ядра у двух видов ацетабулярии: A. mediterranea и A. crenulata, которые различаются формой шляпки. Производилось пересаживание ядра или сращивание стебелька с ризоидом разных видов. Шапочка приобретала форму, присущую тому виду, у которого было взято ядро[29].
При пересадке ядра в A. mediterranea из A. Wettsteini в 1935 году Геммерлинг получал растения по типу Wettsteini, но в некоторых случаях — организмы имели признаки обеих форм. По поводу этих необычных растений он писал[30]:
«Но в 6 случаях сначала возникли образования типа mediterranea (типичные мутовки и возможно недоразвитые шляпы), но затем шляпа Wettsteini. Возникновение образований mediterranea основывается на действии ядра Wettsteini. В этих случаях пересаживалась передняя часть mediterranea, содержавшая уже больше или меньше специфических виду формообразующих веществ. Они пошли в ход в первую очередь и, как и следовало ожидать, индуцировали образования mediterranea; лишь затем подействовало ядро Wettsteini».
Циркадный ритм[править | править код]
Ритм выработки кислорода растением зависит от времени суток: днём оно выделяет больше кислорода, чем ночью, поскольку для реакции фотосинтеза нужен свет. Однако этот ритм сохраняется, если ацетабулярию начать освещать постоянно и круглосуточно: действуют внутренние биологические часы организма. Такие периодические процессы называют циркадианным, или циркадным (околосуточным) ритмом. Они присущи всем живым организмам, клетки которых имеют ядро (включая одноклеточные, растения и грибы, а также животных и человека)[4]:95.
Если поменять ритм освещения (освещать водоросль ночью, а не днём), произойдёт смещение фазы суточного ритма у растения на противоположную. Если после этого поменять ядра у растений, имеющих противоположную фазу, то ритм через короткое время установится по приказу соответствующего ядра[4]:95.
Растение стремится поворачивать свою шляпку так, чтобы на неё падало больше света. Кроме того, хлоропласты перемещаются внутри клетки, чтобы в дневное время оказаться на её поверхности, а ночью часть из них опускаются в её нижнюю часть. Этот ритм внутриклеточного перемещения хлоропластов также сохраняется при постоянной температуре и освещении в течение суток[4]:96.
При неизменных температуре и освещении исследователи измеряли разность электрического потенциала (напряжение) между верхним и нижним концами водоросли, и обнаруживали этот же эндогенный околосуточный ритм[4]:96.
Циркадный ритм сохраняется даже после удаления ядра, и даже после разделения клетки на несколько маленьких частей, что свидетельствует о местонахождении биологических часов в цитоплазме или на наружной мембране клетки[4]:95.
В произведениях культуры[править | править код]
- Жюль Верн в романе «Двадцать тысяч льё под водой» (1869—1870) упоминает ацетабулярии среди прочих водных растений во владениях капитана Немо[31].
Литература[править | править код]
- Куреалов Л. И., Комарницкий Н. Ацетабулярия, Курс низших растений, 3-е изд. — М., 1945, С. 84—86.
Атлас фотографий[править | править код]
- Sigrid Berger. Photo-Atlas of living Dasycladales
- Acetabularia acetabulum (= mediterranea[32]) 19 фото
- Acetabularia caliculus 7 фото
- Acetabularia crenulata 21 фото
- Acetabularia dentata 9 фото
- Acetabularia farlowii 7 фото
- Acetabularia kilneri 9 фото
- Acetabularia major 3 фото
- Acetabularia peniculus 9 фото
- Acetabularia ryukyuensis 10 фото Архивная копия от 21 декабря 2009 на Wayback Machine
- Acetabularia schenckii 15 фото
Примечания[править | править код]
- ↑ Белякова Г. А., Пчелкин А. В. «Водоросли и лишайники». Дата обращения: 30 марта 2010. Архивировано 9 апреля 2009 года.
- ↑ 1 2 3 4 5 6 7 8 9 10 11 12 Ацетабулярия / Н. П. Горбунова // Ангола — Барзас. — М. : Советская энциклопедия, 1970. — (Большая советская энциклопедия : [в 30 т.] / гл. ред. А. М. Прохоров ; 1969—1978, т. 2).
- ↑ 1 2 «Онтогенез», Академия наук СССР. том 19 — 1988 г., стр. 200. Дата обращения: 2 октября 2017. Архивировано 17 октября 2016 года.
- ↑ 1 2 3 4 5 6 7 8 9 10 11 12 Ацетабулярия — водоросль и модель (недоступная ссылка). По материалам журнала «Бильд дер Виссеншафт» (ФРГ) // Наука и жизнь. — 1985. — № 3. — С. 93—96, врезка V.
- ↑ doc. Дата обращения: 23 августа 2010. Архивировано из оригинала 4 марта 2016 года.
- ↑ http://evolution.powernet.ru/library/human_brain.html Архивная копия от 17 ноября 2011 на Wayback Machine копия http://www.peeep.us/cc8c91ef Архивная копия от 8 марта 2016 на Wayback Machine
- ↑ KM.RU Самый крупный одноклеточный организм. 14.10.2004 (недоступная ссылка)
- ↑ Kyle A. Serikawa and Dina F. Mandoli. An analysis of morphogenesis of the reproductive whorl of Acetabularia acetabulum //Planta, Volume 207, Number 1 / Ноябрь 1998 г. pp. 96-104 [1] (недоступная ссылка)
- ↑ 1 2 3 4 5 6 7 8 9 10 11 Морфогенез водоросли Acetabularia Архивировано 1 декабря 2009 года.
- ↑ 1 2 3 Рост растений: первичные механизмы. Валентин Ильич Кефели, Научный совет по проблемам физиологии и биохимии растений (Академия наук СССР), Институт физиологии растений им. К. А. Тимирязева. 1978 — Всего страниц: 288, стр. 38 [2]
- ↑ 1 2 3 4 Курс низших растений. Учебник для студентов. Под ред. М. В. Горленко. — М.: Высшая школа, 1981. — 504 c., страница 144—148 (DJVU)
- ↑ Kwang W. Jeon. International Review of Cytology: A Survey of Cell Biology, Academic press, 1998, стр. 12. [3]
- ↑ Sigrid Berger. Photo-Atlas of living Dasycladales. Systematics and age of the Dasycladales. [4] Архивная копия от 5 января 2010 на Wayback Machine
- ↑ Успехи современной биологии, тома 85-86. Академия наук СССР. стр. 75, 80. [5]
- ↑ 1 2 Николай Петрович Дубинин «Эволюция популяций и радиация» 1966, стр. 51
- ↑ «Наука и жизнь», выпуски 1-4 1985 г. стр. 95
- ↑ см. фото http://paleopolis.rediris.es/cg/CG2006_BOOK_02/CG2006_B02_Fig_099.htm Архивная копия от 21 декабря 2009 на Wayback Machine
- ↑ см. фото http://paleopolis.rediris.es/cg/CG2006_BOOK_02/CG2006_B02_Fig_135.htm Архивная копия от 21 декабря 2009 на Wayback Machine
- ↑ Christiaan Hoek,D. G. Mann,Hans Martin Jahns. Algae: an introduction to phycology. Cambridge univercity press. — 1995, стр. 443. [6]
- ↑ BERGER Sigrid ; LIDDLE Larry B. The life cycle of Acetabularia (Dasycladales, Chlorophyta): textbook accounts are wrong. ISSN : 0031-8884. Phycologia A. 2003, vol. 42, n° 2, pp. 204—207 [4 pages] [7]
- ↑ 1 2 Frederick S. Russell. Advances in marine biology, том 14. стр. 142
- ↑ scinote.ru Исследования клетки | Заметки о науке (недоступная ссылка)
- ↑ Frederick S. Russell. Advances in marine biology, том 14. стр. 142
- ↑ М. А. Грачев, Лимнологический институт СО РАН (Иркутск), «Наука из первых рук», материалы конференции, 2006 ISSN 1810-3960.[8] Архивная копия от 12 июня 2016 на Wayback Machine
- ↑ Günter Vogel und Hartmut Angermann: Taschenbuch der Biologie. 2. Auflage. VEB Gustav Fischer Verlag, Jena 1979, Lizenznummer:261 700/174/79 Bd.1
- ↑ Dr.rer.nat Erik Heinz Benedix, Dr.rer.nat.habil S. Jost Casper u. a.: Urania Pflanzenreich. 1. Auflage. Urania-Verlag, Leipzig/Jena/Berlin 1974, VLN. 212—475/26/74 Bd.: Niedere Pflanzen
- ↑ Wolfgang Miram und Karl-Heinz Scharf (Hrsg.): Biologie heute SII. Schroedel Schulbuchverlag, Hannover 1988, ISBN 3-507-10540-3
- ↑ «Наука в сибири» N 45 (2630) 22 ноября 2007 г. Дата обращения: 30 марта 2010. Архивировано 6 июля 2013 года.
- ↑ Hammerling, 1943 в кн. Srb et al., 1965, цит. по Жимулев Игорь Федорович. Общая и молекулярная генетика. Курс лекций для студентов 3-го курса. Глава 14. Генетика развития. Архивированная копия. Дата обращения: 30 марта 2010. Архивировано 19 апреля 2009 года.
- ↑ Геммерлинг, 1935. цит. по И. Е. Глущенко «Вегетативная гибридизация растений» (1948 г.), стр. 24-25. Архивированная копия. Дата обращения: 9 сентября 2010. Архивировано 9 сентября 2010 года.
- ↑ Жюль Верн. Двадцать тысяч льё под водой. Пер. с фр. — Н.Яковлева, Е.Корш. «Собрание сочинений», т.4. М., Государственное издательство художественной литературы, 1956. [9] Архивная копия от 16 марта 2010 на Wayback Machine
- ↑ Refdoc